LA NEURONA Y SU FUNCIONAMIENTO

INSTITUTO DE ESTUDIOS UNIVERSITARIOS
STEPHANIE PAMELA PACHECO JIMÉNEZ
MATRICULA: 137910
GRUPO: LP49
FUNDAMENTOS BIOLÓGICOS DEL COMPORTAMIENTO
MTRA.: MARÍA ALEJANDRA MORAN MAYEN.
MODULO 2: ACTIVIDAD DE APRENDIZAJE 2
LA NEURONA Y SU FUNCIONAMIENTO
BLOG
ATITALQUIA HIDALGO, A 2 DE DICIEMBRE DE 2020
Tu capacidad de percibir tu entorno, de ver, oír y oler lo que te rodea, depende de tu sistema
nervioso; también tu habilidad para reconocer dónde estás y recordar si has estado allí
antes. De hecho, ¡tu mera capacidad de preguntarte dónde estás depende de tu sistema
nervioso!
Todos estos procesos dependen de las células interconectadas que forman el sistema
nervioso. Como el corazón, los pulmones y el estómago, el sistema nervioso se compone de
células especializadas. Estas incluyen células nerviosas (o neuronas) y células
gliales (o glía). Las neuronas son las unidades funcionales básicas del sistema nervioso y
generan señales eléctricas llamadas potenciales de acción que les permiten transmitir
información rápidamente a largas distancias. La glía también es esencial para la función del
sistema nervioso, pero su principal función es apoyar a las neuronas.
Anatomía de una neurona
Las dendritas

Los axones
- Las dendritas tienden adelgazarse conforme se alargan y suelen estar cubiertas de pequeños bultos llamados espinas. En contraste, el axón suele conservar el mismo diámetro en la mayor parte de su longitud y no tiene espinas.
- El axón surge del cuerpo celular en un área especializada llamada cono axónico. En neuronas motoras e interneuronas, es ahí donde inicia el potencial de acción.
- Por último, muchos axones están cubiertos con una sustancia aislante especial llamada mielina, que les ayuda a transmitir rápidamente los impulsos nerviosos. La mielina nunca se encuentra en dendritas.
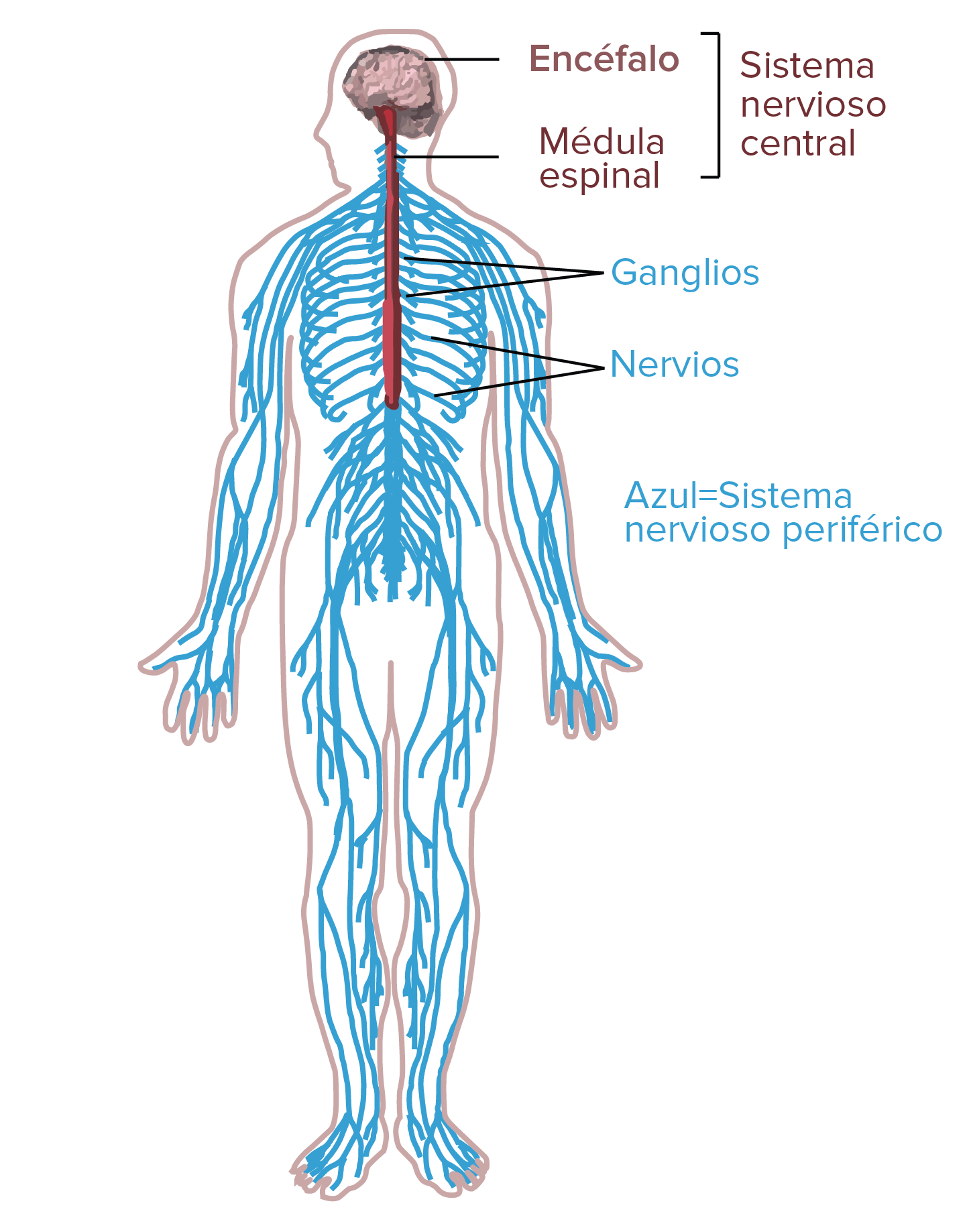
El sistema nervioso humano
- El sistema nervioso central (SNC) consiste del encéfalo y la médula espinal. En el SNC es donde ocurre todo el análisis de la información.
- El sistema nervioso periférico (SNP), compuesto por las neuronas y partes de las neuronas que se encuentran fuera del SNC, incluye neuronas sensoriales y neuronas motoras. Las neuronas sensoriales llevan señales hacia el SNC y las neuronas motoras llevan señales enviadas por el SNC.
Tipos de neuronas
Neuronas sensoriales
Neuronas motoras
Interneuronas
Las funciones básicas de una neurona
- Recibir señales (o información).
- Integrar las señales recibidas (para determinar si la información debe o no ser transmitida).
- Comunicar señales a células blanco (músculos, glándulas u otras neuronas).
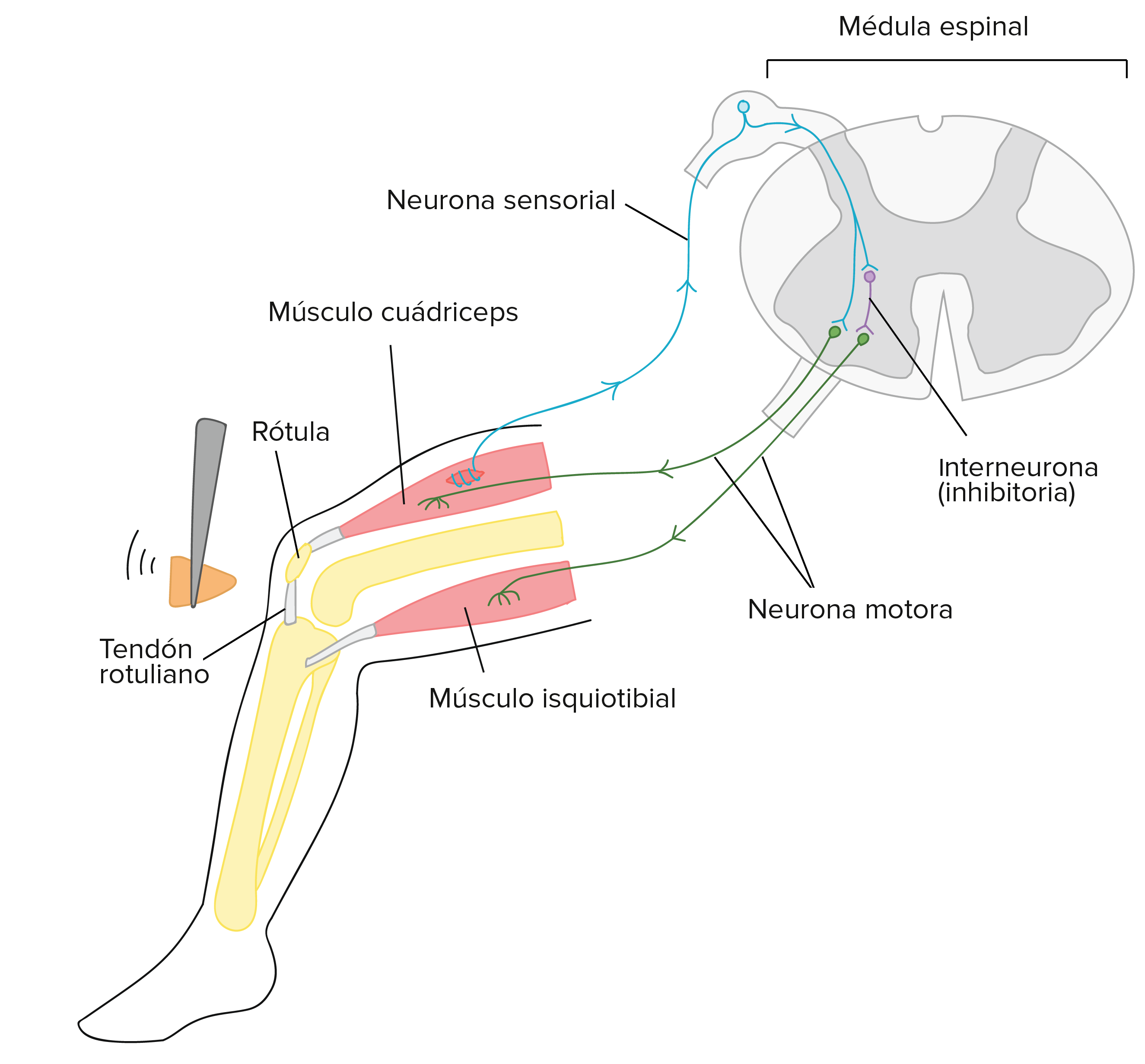
El reflejo rotuliano
Células gliales
Al principio de este artículo, dijimos que el sistema nervioso se compone de dos tipos de células: neuronas y glía; siendo las neuronas las que actúan como unidad funcional básica del sistema nervioso, y las gliales las que desempeña un papel secundario de apoyo. Así como los actores secundarios son esenciales para el éxito de una película, la glía es esencial para la función del sistema nervioso. De hecho, hay muchas más células gliales en el encéfalo que neuronas.
Hay cuatro tipos principales de células gliales en el sistema nervioso vertebrado adulto. Tres de estos —astrocitos, oligodendrocitos y microglía— solo se encuentran en el sistema nervioso central (SNC). El cuarto tipo, las células de Schwann, solo se encuentra en sistema nervioso periférico (SNP).
Tipos de glía y sus funciones
Los astrocitos son el tipo de células gliales más numeroso. De hecho, ¡son las células más numerosas en el encéfalo! Hay astrocitos de diferentes tipos y tienen varias funciones. Ayudan a regular el flujo de sangre en el encéfalo, mantienen la composición del líquido que rodea las neuronas y regulan la comunicación entre las neuronas en la sinapsis. Durante el desarrollo, los astrocitos ayudan a que las neuronas lleguen a sus destinos y contribuyen a la formación de la barrera hematoencefálica, que ayuda a aislar el encéfalo de sustancias potencialmente tóxicas en la sangre.
La microglía está relacionada con los macrófagos del sistema inmunitario y actúan como carroñeros que eliminan células muertas y otros residuos.
Los oligodendrocitos del SNC y las células de Schwann del SNP comparten una función similar. Ambos tipos de células gliales producen mielina, la sustancia aislante que forma una funda alrededor de los axones de muchas neuronas. La mielina aumenta dramáticamente la velocidad con la que un potencial de acción viaja por el axón y desempeña un papel crucial en la función del sistema nervioso.


- Si el potencial de membrana se vuelve más positivo que el potencial de reposo, se dice que la membrana se despolariza.
- Si el potencial de membrana se vuelve más negativo que el potencial de reposo, se dice que la membrana se hiperpolariza.
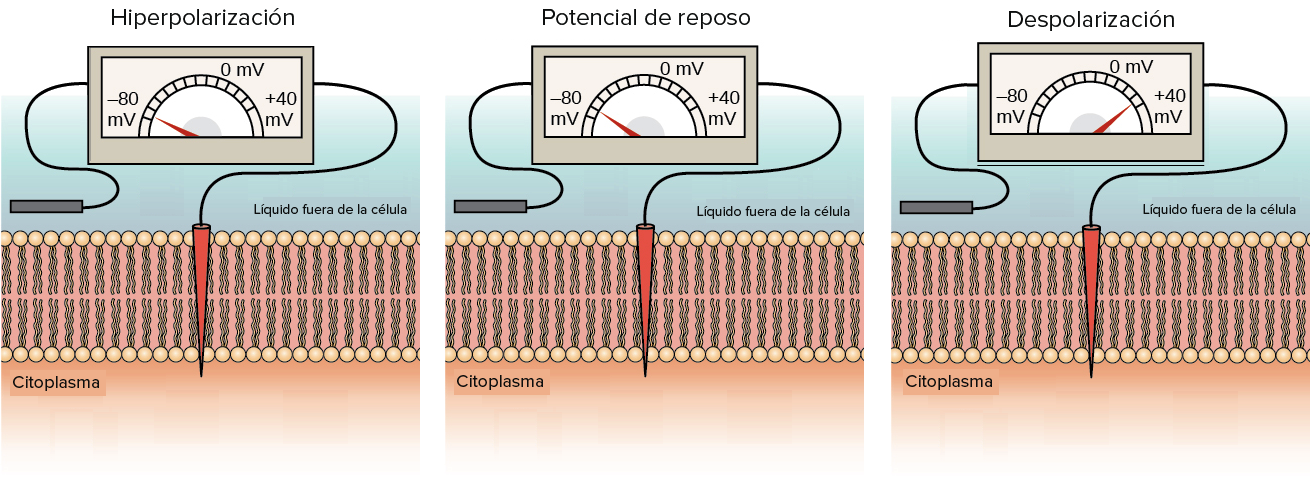
¿De dónde proviene el potencial de membrana en reposo?
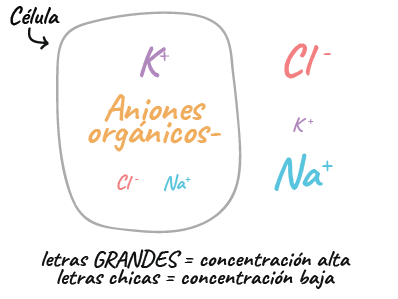
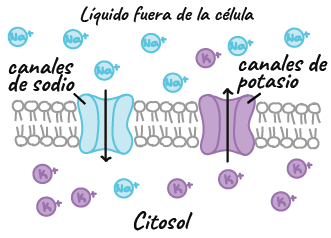
Tipos de iones que se encuentran en las neuronas
- Iones con carga positiva (cationes): sodio (start text, N, a, end text, start superscript, plus, end superscript) y potasio (start text, K, end text, start superscript, plus, end superscript)
- Iones con carga negativa (aniones): cloruro (start text, C, l, end text, start superscript, minus, end superscript) y aniones orgánicos.
Cómo los iones cruzan la membrana
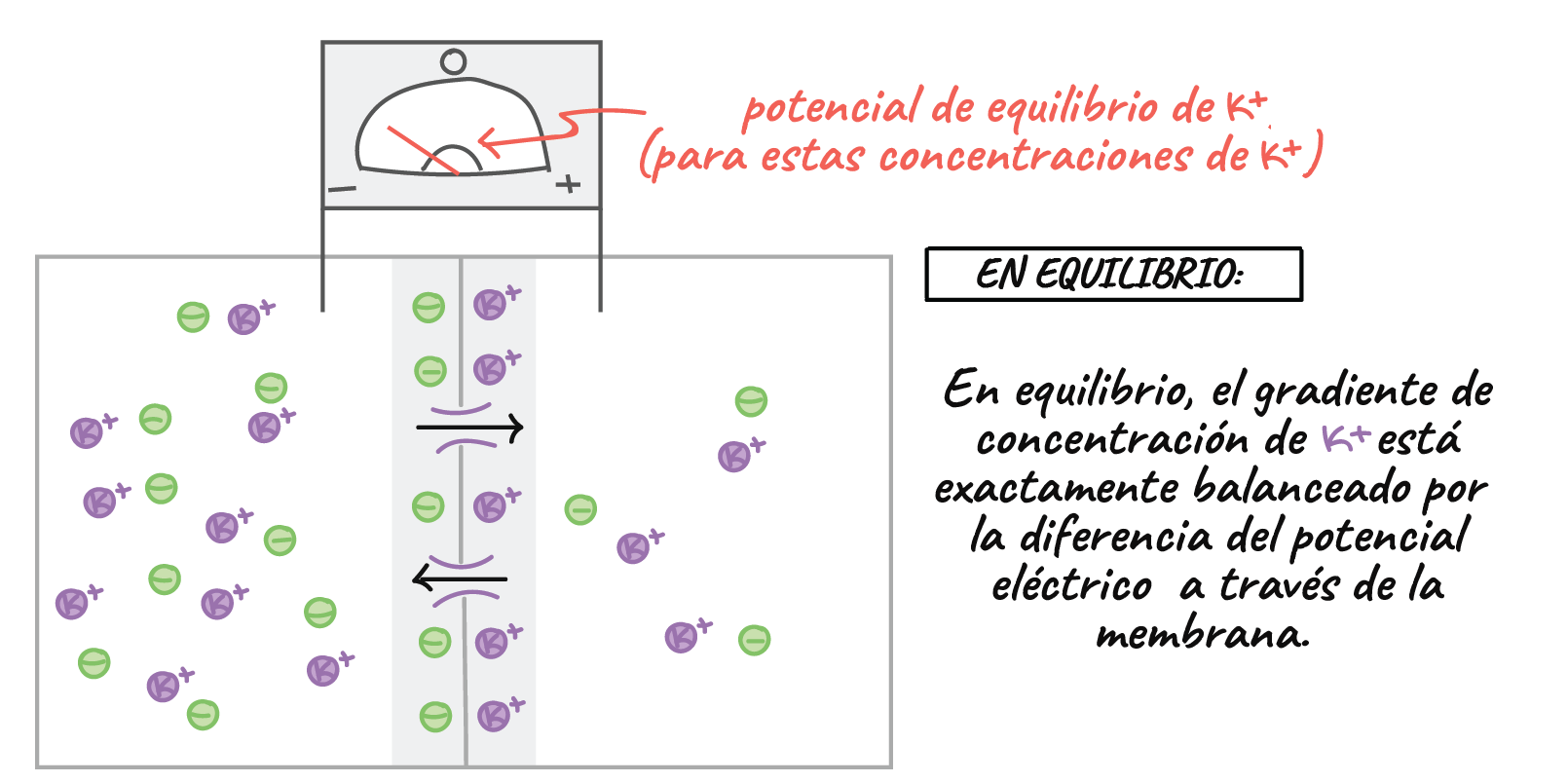
El potencial de equilibrio
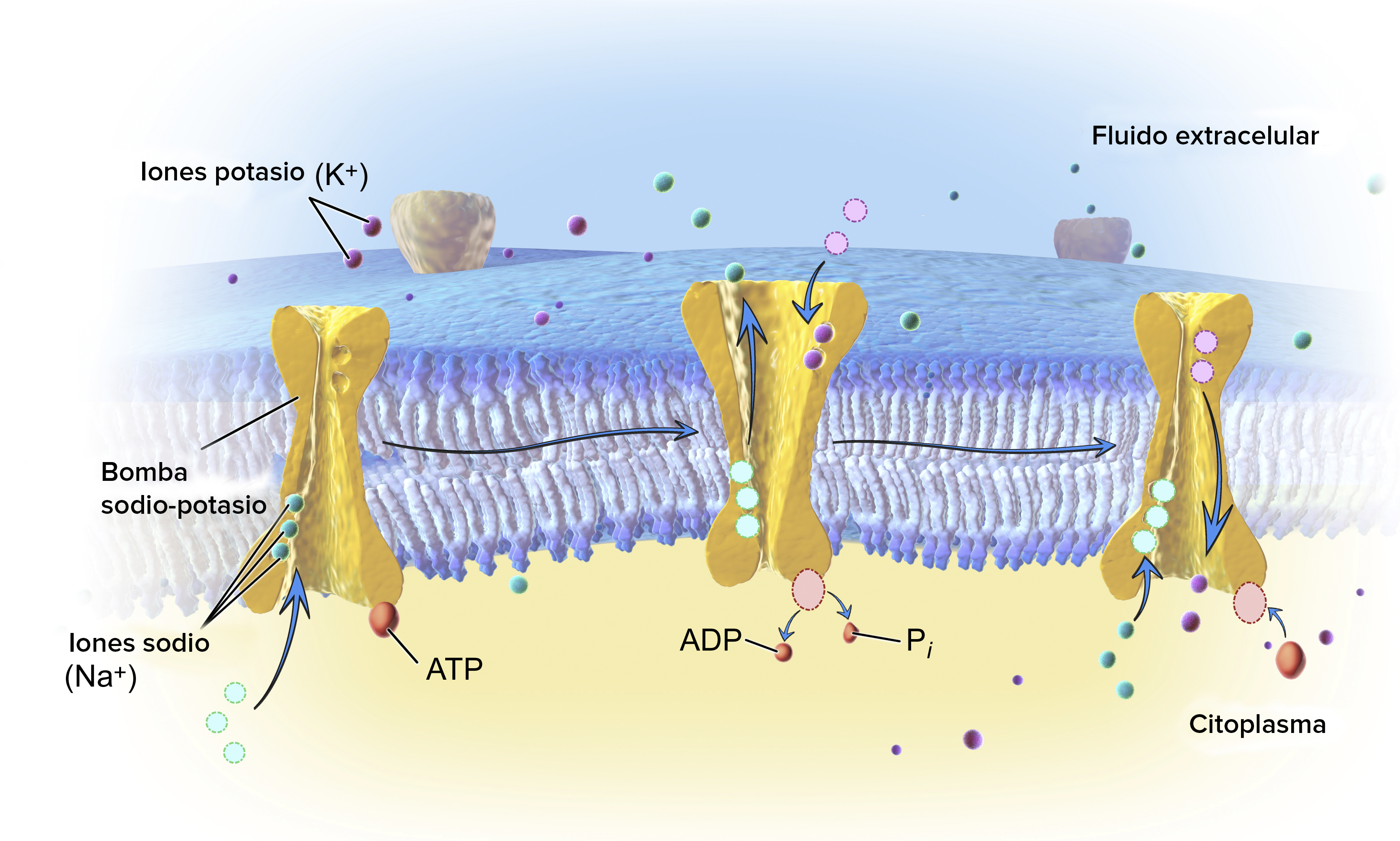
La bomba de start text, N, a, end text, start superscript, plus, end superscript-start text, K, end text, start superscript, plus, end superscript mantiene los gradientes de start text, N, a, end text, start superscript, plus, end superscript y start text, K, end text, start superscript, plus, end superscript
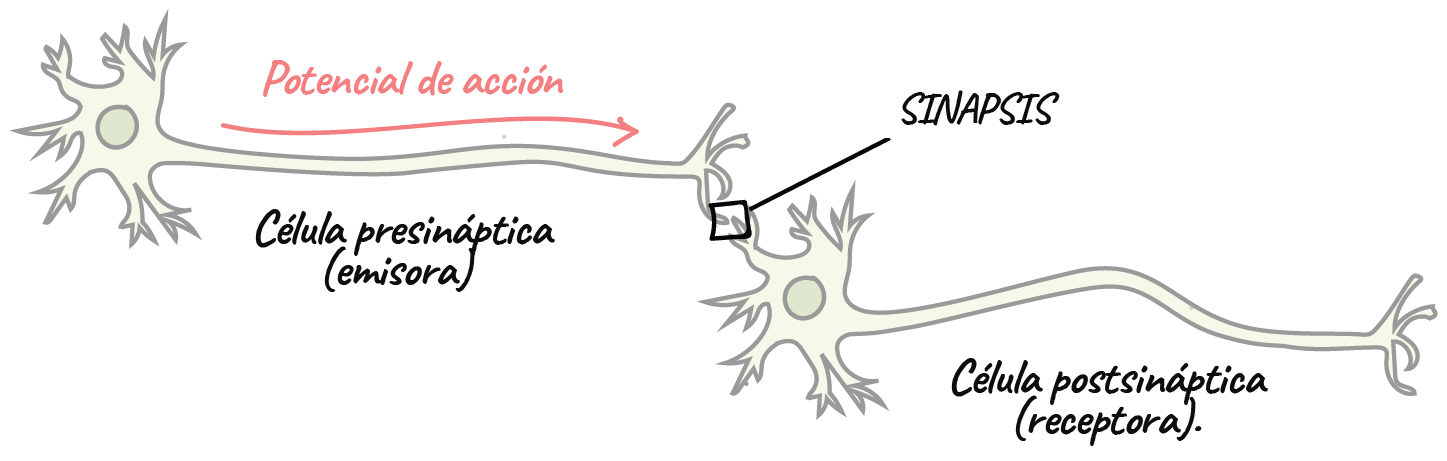
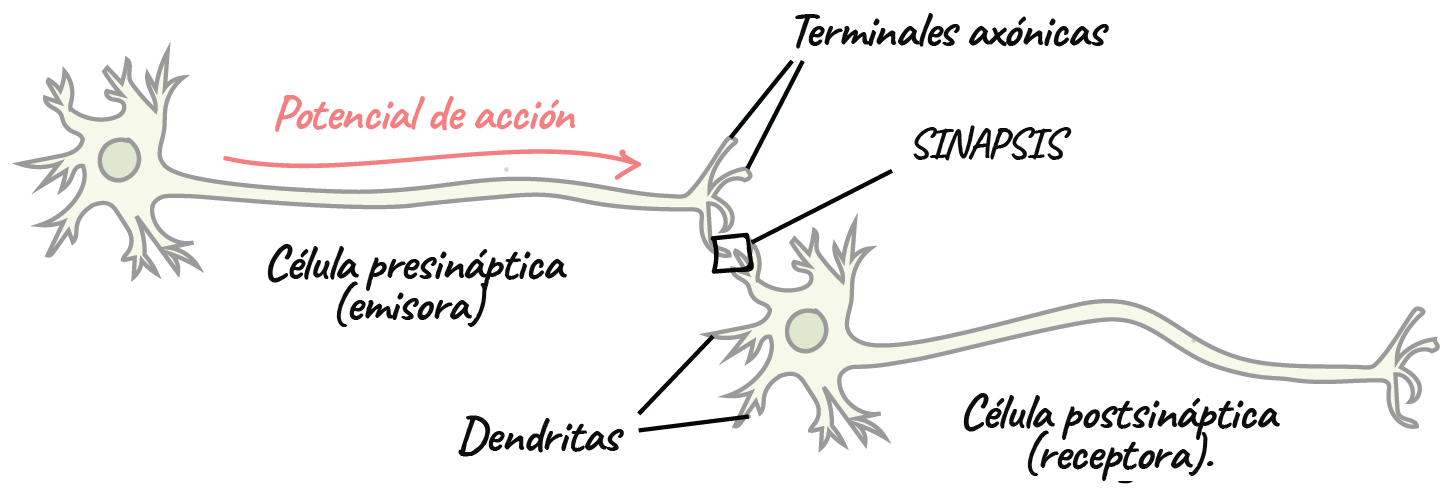
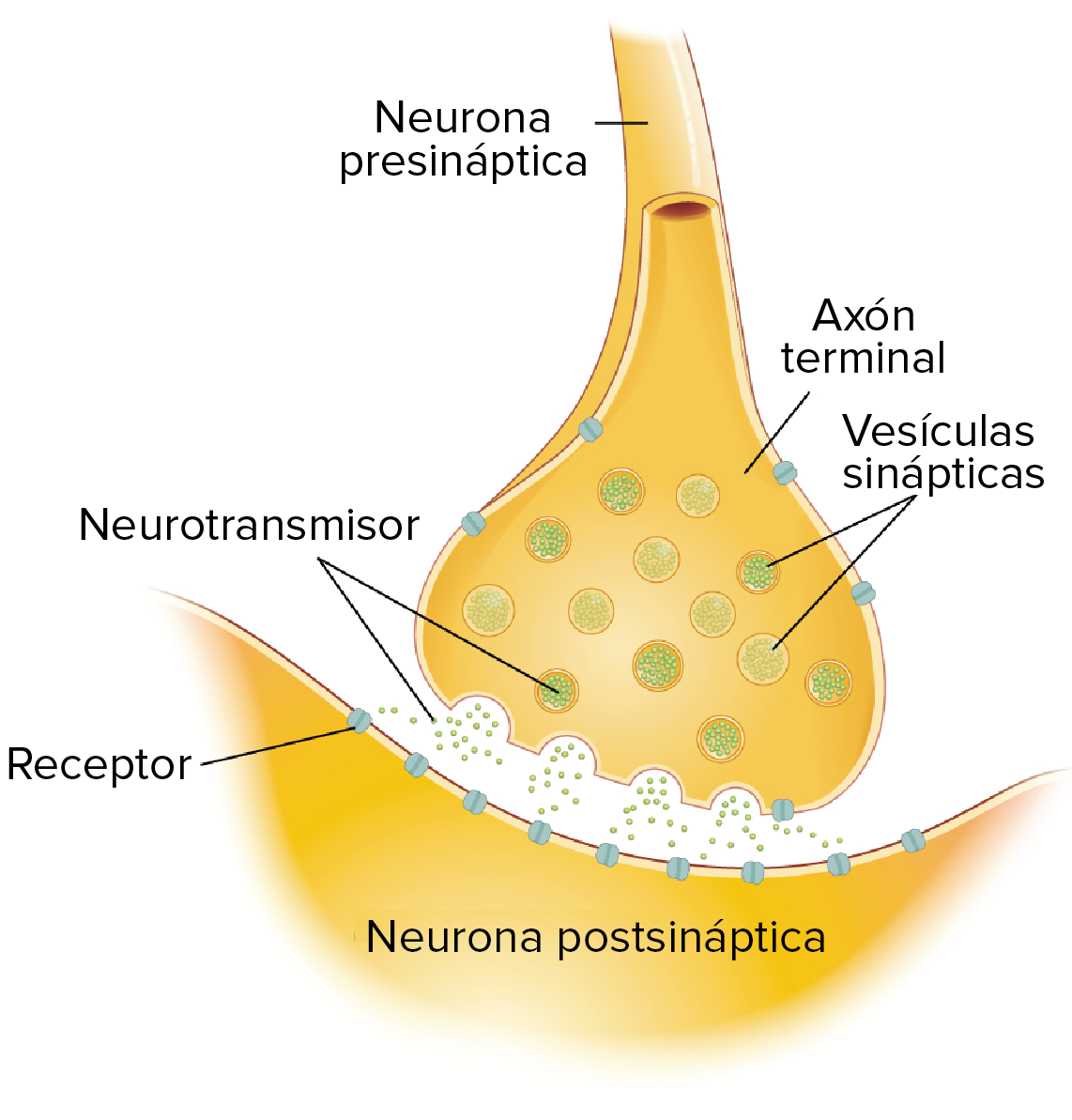
La sinapsis
- Las neuronas se comunican entre sí en las uniones llamadas sinapsis. En una sinapsis, una neurona envía un mensaje a una neurona blanco: otra célula.
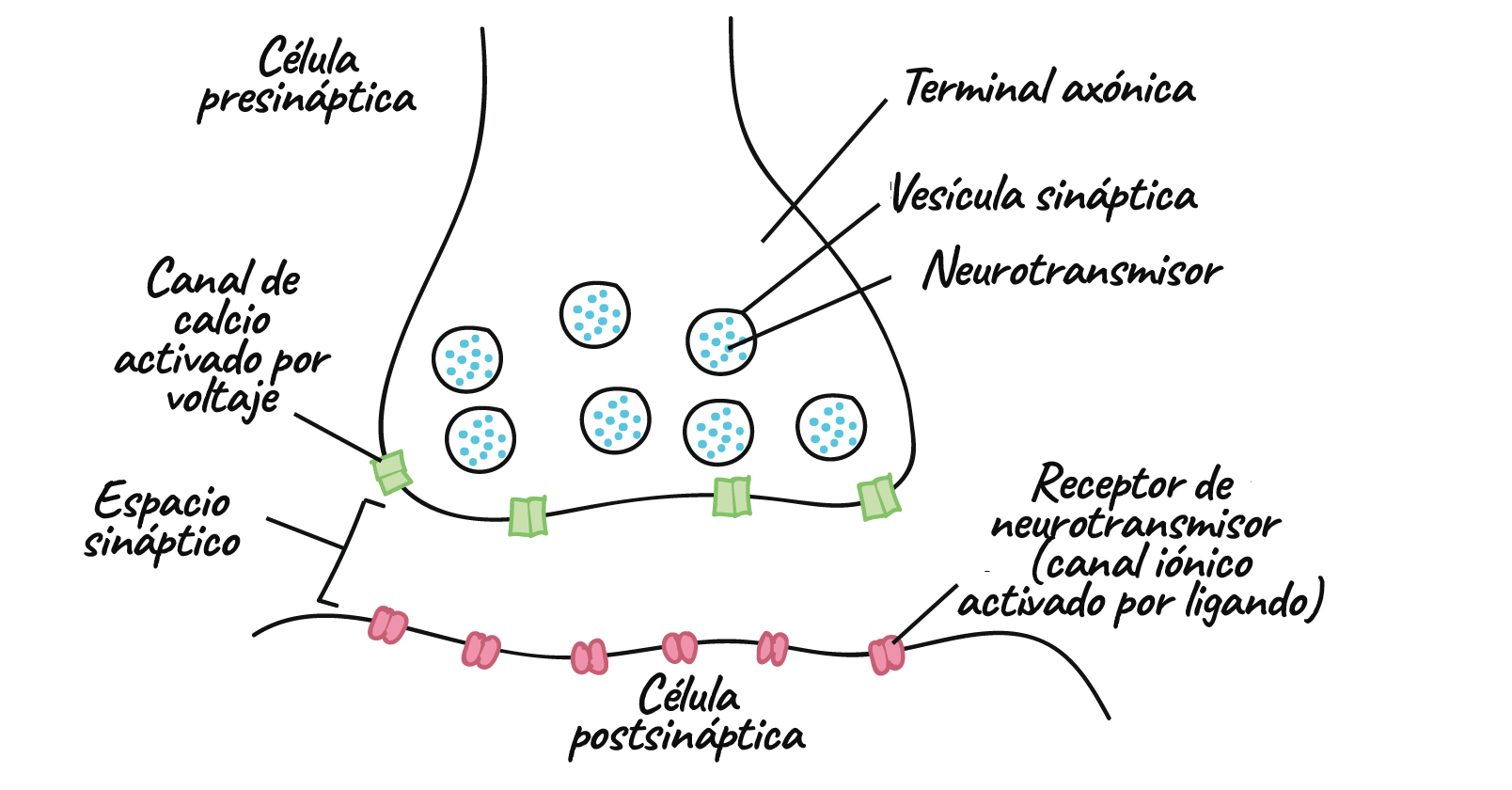
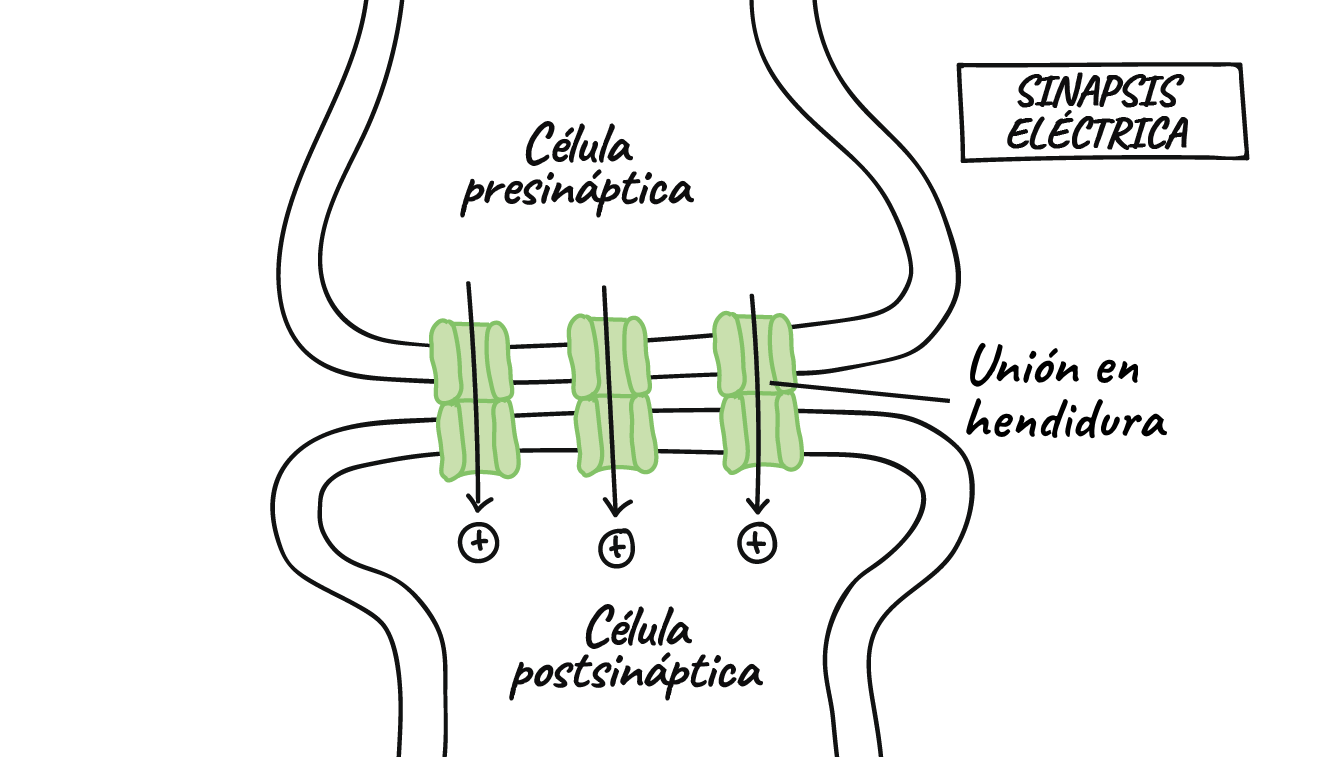
- La mayoría de las sinapsis son químicas, las cuales se comunican con mensajeros químicos. Otras sinapsis son eléctricas, en ellas los iones fluyen directamente entre células.
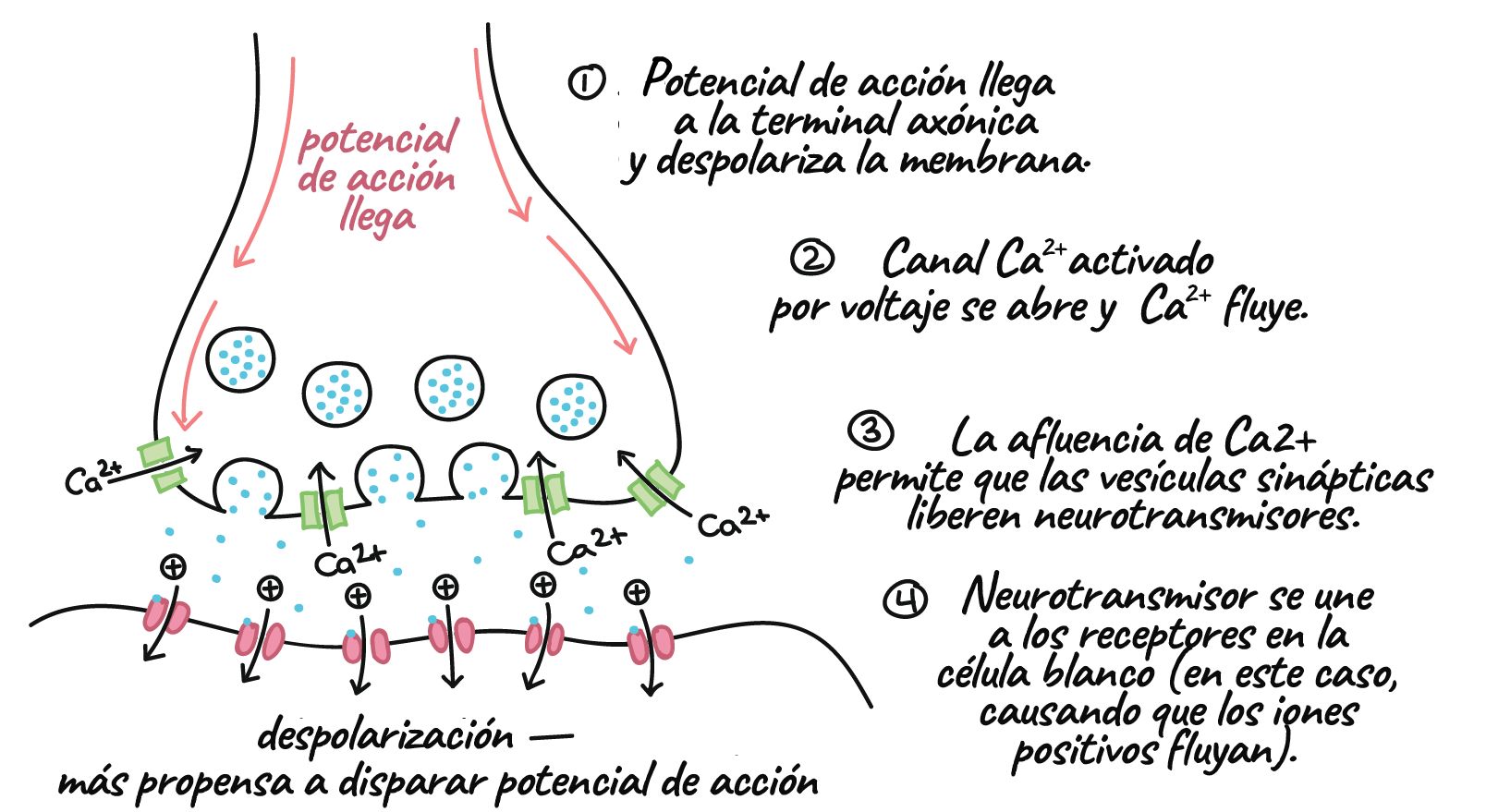
- En una sinapsis química, un potencial de acción provoca que la neurona presináptica libere neurotransmisores. Estas moléculas se unen a receptores en la célula postsináptica y modifican la probabilidad de que esta dispare un potencial de acción.
Transmisión en las sinapsis químicas
Potenciales postsinápticos excitatorios e inhibitorios
- En algunos casos, el cambio provoca que la célula blanco sea más propensa a disparar su propio potencial de acción. En este caso, el cambio en el potencial de membrana se llama potencial excitatorio postsináptico o PEPS.
- En otros casos, el cambio provoca que la célula blanco sea menos propensa a disparar su propio potencial de acción y se llama potencial inhibitorio postsináptico o PIPS.
Sinapsis eléctricas
Neurotransmisores: convencionales y no convencionales
Neurotransmisores convencionales
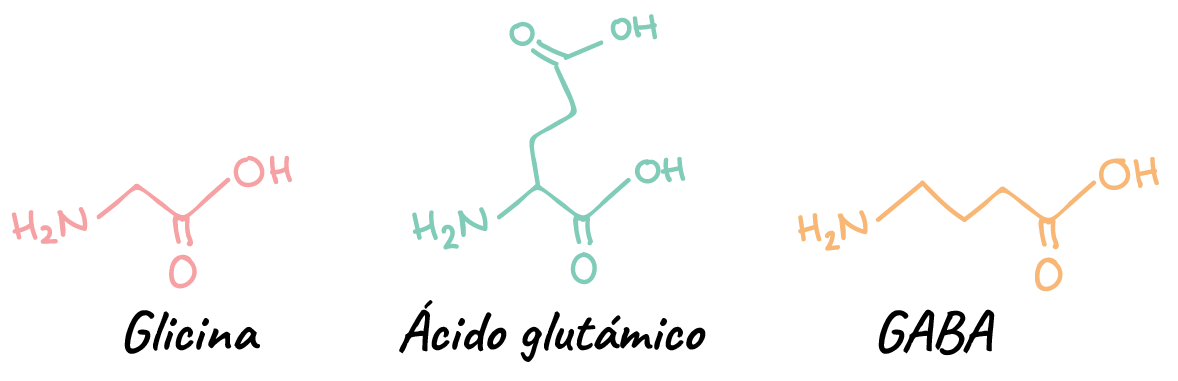
Neurotransmisores de molécula pequeña
- Los aminoácidos neurotransmisores glutamato, GABA (ácido γ-aminobutírico) y glicina. Todos estos son aminoácidos, aunque GABA no es un aminoácido que se encuentre en las proteínas.
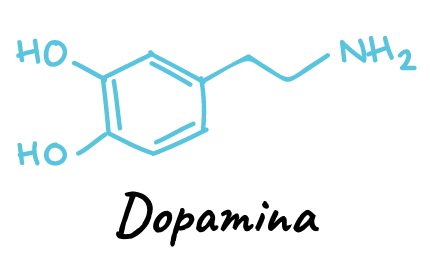
- Las aminas biogénicas dopamina, norepinefrina, epinefrina, serotonina e histamina, que se hacen a partir de aminoácidos precursores.
- Los neurotransmisores purinérgicos ATP y adenosina, que son nucleótidos y nucleósidos.

- La acetilcolina, que no encaja en ninguna de las otras categorías estructurales, pero es un neurotransmisor clave en las uniones neuromusculares (donde se conectan los nervios a los músculos), así como en algunas otras sinapsis.



Neuropéptidos

Hanani, M. (2005). Satellite glial cells in sensory ganglia: From form to function (Células gliales satélite en los ganglios sensoriales: de forma a función). Brain Research Reviews, 48, 457-476. http://dx.doi.org/10.1016/j.brainresrev.2004.09.001. Consultado en http://cc.oulu.fi/~aheape/GW_article_8.pdf.Kandel, E. R., Schwartz, J. H y Jessell, T. M. (1995). Nerve cells and behavior (Células nerviosas y comportamiento). En Essentials of neuroscience and behavior (pp. 21-31). Norwalk, CT: Appleton & Lange.Nicholls, J. G., Martin, A. R., Wallace, B. G y Fuchs, P. A. (2001). Principles of signaling and organization (Principios de señalización). En From neuron to brain (4ta ed., pp. 3-9). Sunderland, MA: Sinauer Associates.OpenStax College, Biología. (29 de septiembre de 2015). Neurons and glia (Neuronas y glía). En OpenStax CNX. Consultado en http://cnx.org/contents/GFy_h8cu@9.87:c9j4p0aj@3/Neurons-and-Glial-Cells.Purves, D., Augustine, G. J., Fitzpatrick, D., Katz, L. C., LaMantia, A.-S y McNamara, J. O. (1997). The organization of the nervous system (La organización del sistema nervioso). En Neuroscience (pp. 1-10). Sunderland, MA: Sinauer Associates.Reece, J. B., Urry, L. A., Cain, M. L., Wasserman, S. A., Minorsky, P. V y Jackson, R. B. (2011). Nervous systems consist of circuits of neurons and supporting cells (El sistema nervioso consiste de circuitos de neuronas y de células de apoyo). En Campbell biology (10° ed., pág. 1080-1084). San Francisco, CA: Pearson.Reece, J. B., Urry, L. A., Cain, M. L., Wasserman, S. A., Minorsky, P. V y Jackson, R. B. (2011). Neuron structure and organization reflect function in information transfer (La estructura neuronal y su organización reflejan su función en la transferencia de información). En Campbell biology (10° ed., pág. 1062-1064). San Francisco, CA: Pearson.Sadava, D. E., Hillis, D. M., Heller, H. C y Berenbaum, M. R. (2009). Neurons and nervous systems (Las neuronas y el sistema nervioso). En Life: The science of biology (9na ed., pp. 988-993). Sunderland, MA: Sinauer Associates.Sadava, D. E., Hillis, D. M., Heller, H. C y Berenbaum, M. R. (2009). The spinal cord transmits and processes information (La médula espinal transmite y procesa información). En Life: The science of biology (9na ed., pp. 1033-1034). Sunderland, MA: Sinauer Associates.Solomon, E. P. (2016). Sensory receptors (Receptores sensoriales). En Introduction to human anatomy and physiology (4ta ed., pp. 145). Maryland Heights, MO: Saunders.Swift, A. (2015). Pain management 2: Transmission of pain signals to the brain (Tratamiento del dolor 2: transmisión de señales de dolor hacia el cerebro). Nursing Times, 111(40), 22-26. Consultado en http://www.nursingtimes.net/clinical-subjects/pain-management/pain-management-2-transmission-of-pain-signals-to-the-brain/5090685.fullarticle.
CONCLUSIÓN:
Conocer la estructura y funcionamiento de la neurona como base del estudio de la conducta humana es interesante si tomamos en cuenta que esencialmente constituye nuestro cerebro y nuestro sistema nervioso y que es responsable de los pensamientos y de nuestros sentimientos y tal vez de todos nuestros sentidos yo creo que fácilmente estaría entre el segundo o en el primer puesto en cuánto a importancia. Es entonces que se entiende que el comportamiento también puede definirse como un cambio en la actividad de un organismo en respuesta a un estímulo, una señal externa o interna, o una combinación de señales y que para entender completamente un comportamiento, queremos saber qué lo provoca, cómo se desarrolla en un individuo, cómo beneficia a un organismo y cómo evoluciona. Algunos comportamientos son innatos, o programados genéticamente, mientras que otros son aprendidos, o desarrollados por la experiencia. En muchos casos, los comportamientos tienen un componente innato y un componente aprendido y todo esto nos quedará mas claro si aprendemos lo necesario al funcionamiento de las neuronas.
Comentarios
Publicar un comentario